Rudimente – Belege für Makroevolution?
Ein beliebter Beleg von Evolutionsvertretern für Makroevolution (Evolution von der Urzelle bis zum Menschen) sind sogenannte Rudimente. Mit Rudimenten sind Organe und Strukturen gemeint, die Überreste der Evolution sein sollen. Sie sollen ihre ursprüngliche Funktion völlig oder zum Teil verloren oder ihre Funktion gewechselt haben.
Oft haben sich jedoch anfangs als rudimentär angenommene Organe als reine Wissenslücken erwiesen, deren Funktion später erkannt wurde. Bevor wir jedoch konkret zu einigen angeblichen Rudimenten kommen, soll geklärt werden, inwiefern Rudimente der Schöpfungslehre tatsächlich widersprechen würden:
Naturalisten nehmen an, Rudimente seien nur im Rahmen der Evolutionstheorie erklärbar. Doch dem ist nicht so. Auch im Schöpfungsmodell sind Rudimente denkbar: Auf Grundlage des im Schöpfungsberichtes enthaltenen Sündenfalls ist es zu erwarten, dass die anfangs perfekte Schöpfung beginnt zu degenerieren. In dem Rahmen ist es durchaus möglich, dass Organe, die nicht zwingend notwendig zum Überleben sind, ihre Funktion zum Teil oder ganz verlieren.
Liste der behandelten Rudimente:
- Der rückläufige Kehlkopfnerv
- Der Wurmfortsatz des Blinddarms
- Die Weisheitszähne
- Das Steißbein
- Die Abdominalknochen der Wale
- Die menschliche Körperbehaarung
- Darwins Ohrhöcker
- Flugunfähige Vögel
- Hervorstehende Eckzähne
- Das Nickhaut-Rudiment
- Augenreste bei Höhlenfischen
- Die Afterklauen der Python und Boa
- Blinde Maulwürfe
- Der Greifreflex
- Die männlichen Brustwarzen
- Schulter- und Beckenknochenreste bei Blindschleichen
1. Der rückläufige Kehlkopfnerv:
Der rückläufige Kehlkopfnerv macht einen scheinbar nutzlosen Umweg über das Herz:
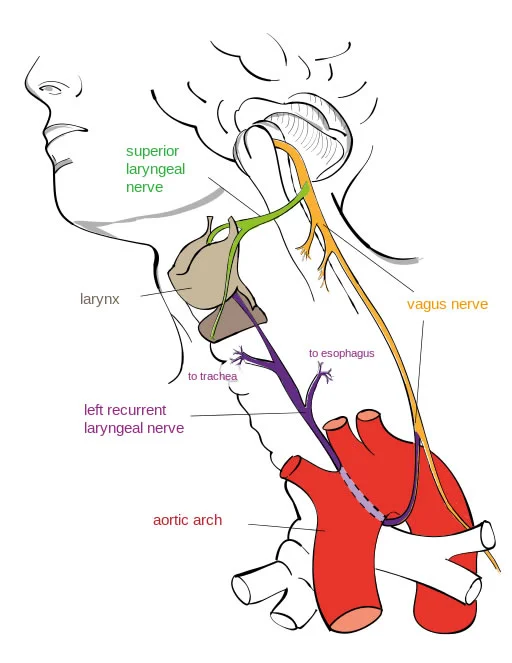
Evolutionsvertreter sehen in diesem Umweg keinen Nutzen und die einzige Erklärung in der hypothetischen Entwicklung vom Fisch zum Säugetier, bei der der Hals länger geworden sei und das Herz tiefer in den Körper gewandert sein soll.
Doch ist inzwischen nachgewiesen, dass dieser Umweg keineswegs nutzlos ist.
Nerven leiten bekanntlich nicht nur Befehle vom Gehirn zu den Organen, sondern auch sensorische und viszerale Informationen von den Organen zum Gehirn. Sie geben dem Gehirn sozusagen „Rückmeldung“. Das ist wichtig, damit das Gehirn in jeder Situation die richtigen Befehle geben kann.
Neben seiner Beteiligung an der Sprachproduktion reguliert der Kehlkopf auch den Zustrom von Luft in die Luftröhre und von Nahrung und Flüssigkeiten in die Speiseröhre. Das ist durchaus eine lebenswichtige Aufgabe, wenn man etwa bedenkt, dass eine mit Nahrung verstopfte Luftröhre zum Erstickungstod führen kann. Anatomisch (vom Aufbau und der Anordnung her) ist es offensichtlich, dass der rückläufige Kehlkopfnerv auf seinem „Umweg“ zahlreiche autonome und sensorische Nerven versorgt, die sich zur Luftröhre, Speiseröhre, der Aorta und zum Herzen verzweigen.1 Somit liegt dieser scheinbare Umweg genau zwischen den Organen, von denen der rückläufige Kehlkopfnerv Informationen für das Gehirn sammelt und erfüllt somit einen Zweck.
In sehr seltenen Fällen werden Menschen geboren, die diesen Umweg nicht haben. Tatsächlich wurden bei manchen von ihnen leichte Schluckbeschwerden und ein Fremdkörpergefühl beim Schlucken festgestellt.2 Das ist ein zusätzliches Indiz für die Funktion des rückläufigen Kehlkopfnerves.
Der rückläufige Kehlkopfnerv stellt also absolut kein Problem für einen Schöpfer dar. Die Behauptung, der Umweg sei im Lauf der Evolution entstanden, ist lediglich eine Interpretation unter der vorausgesetzten Annahme, dass es Makroevolution gegeben hat. Diese Interpretation mag zwar meines Wissens nicht widerlegt werden können, aber sie kann auch nicht eindeutig belegt werden. Letztlich scheint der rückläufige Kehlkopfnerv mit beiden Modellen vereinbar zu sein.
Ausführlicher wird der rückläufige Kehlkopfnerv hier behandelt.
2. Der Wurmfortsatz des Blinddarms:
Ein ebenfalls sehr bekanntes und längst widerlegtes angebliches Rudiment ist der Wurmfortsatz des Blinddarms. Früher dachte man dieser sei vollkommen nutzlos und früher ein funktionstüchtiger Teil des Blinddarms gewesen, der später aber durch geänderte Essgewohnheiten nicht mehr gebraucht wurde und sich zurück entwickelte.
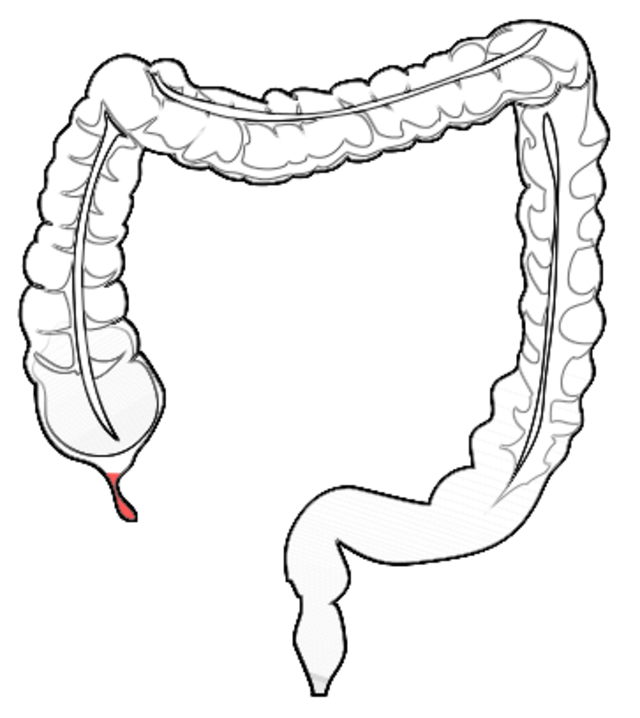
Doch inzwischen ist längst bekannt, dass dieser Wurmfortsatz wichtige Funktionen erfüllt. Er hat eine ähnliche Funktion wie die Mandeln, indem er Krankheitserreger abwehrt. Außerdem dient er als Zufluchtsstätte für symbiontische Bakterien, die das Wachstum nützlicher Darmbakterien fördern.
Wird der Darm durch Durchfall entleert, bleiben diese Bakterien verschont und ermöglichen ein schnelles Wiederbesiedeln des Darms. Das ist wichtig, da eine gesunde Darmflora besonders für die Individualentwicklung (persönliche Entwicklung während der Kindheit) wichtig ist.
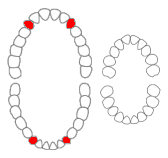
Unabhängig von der Funktion stellt der Wurmfortsatz in anderer Hinsicht ein Problem für die Evolutionstheorie (ET) dar: Der Wurmfortsatz kommt bei vielen verschiedenen Tierarten vor, die auch aus evolutionstheorethischer Sicht nicht nah verwandt sind. Im Normalfall werden gleiche / ähnliche Merkmale bei verschiedenen Tierarten auf eine gemeinsame Abstammung zurück geführt. Diese werden dann als „homolog“ bezeichnet.
Doch ergeben sich bei Stammbaumrekonstruktionen auf Grundlage morphologischer (äußerlich sichtbarer) Merkmale oft widersprüchliche Stammbäume. Um solche Widersprüche zu lösen, muss angenommen werden, dass ein Teil der Merkmale konvergent (mehrfach unabhängig voneinander) entstanden sind. Im Fall vom Wurmfortsatz kommt er bei mindestens 50 Tierarten vor und müsste mindestens 32 Mal konvergent entstanden sein. Diese Grafik veranschaulicht das:
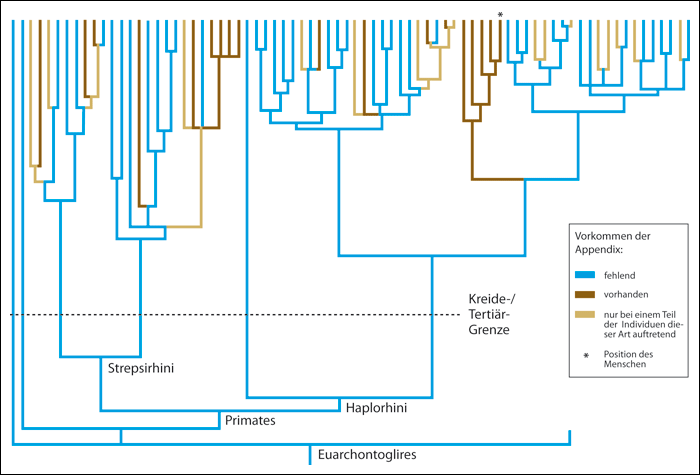
Das Problem für die ET dabei ist, dass Konvergenzen aus evolutionstheoretischer Sicht äußerst selten sein müssten (Das war früher auch die Annahme, bevor bekannt wurde, wie häufig Konvergenzen sind.), da Evolution ungerichtet verläuft und nicht zu erwarten ist, dass sie zwei Mal zum gleichen Ergebnis führt. Aber eben das müsste beim Wurmfortsatz und vielen anderen konvergenten Merkmalen der Fall sein, wenn sie tatsächlich evolviert wären.
Die bessere Erklärung ist ein gemeinsamer Schöpfer, der ähnliche Merkmale beliebig oft einbauen und kombinieren kann (Baukastensystem), weil sie einfach gut funktionieren.
Der Wurmfortsatz des Blinddarms ist also problemlos mit einem Schöpfer vereinbar und stellt im Gegenteil sogar noch ein Problem für die ET dar.
Ein ausführlicherer Artikel zum Wurmfortsatz findet sich hier.
3. Die Weisheitszähne:
Fast jeder kennt sie: Zähne, die in unserem Kiefer ausbrechen und für die wir keinen Platz haben. Meistens müssen sie operativ entfernt werden. Tatsächlich handelt es sich hier um eine Art Rudiment. – in dem Sinn, dass zumindest Menschen in wohlhabenden Ländern keine Verwendung mehr für sie haben.
Doch ist das kein Beleg für Makroevolution, denn Probleme bereiten die Weisheitszähne erst seit der industriellen Revolution. Davor gab es wenige bis keine Probleme mit den Weisheitszähnen.3
Die wahrscheinlichste Erklärung ist die unterschiedliche Ernährung: Vor 1700 war die Nahrung im Allgemeinen viel gröber und die Abnutzung der Zähne somit größer. Die größere Abnutzung führte zu einer Verkleinerung der Höhe der Zahnkrone.4
Wenn diese Verkleinerung weit genug fortgeschritten war, schaffte sie mehr Platz im Zahnbogen, der es den Weisheitszähnen ermöglichte, normal auszubrechen und wie jeder andere Backenzahn zu funktionieren.
Schwerer zu kauende Nahrung hat außerdem den Effekt, dass die Kieferkochen größer und robuster werden. Durch die Abnutzung der Zähne und das daraus resultierende Wachstum sowie die Entwicklung der Kiefer wird ein größerer Zahnraum für alle Backenzähne geschaffen.
Dadurch sind die Weisheitszähne nicht nur nützlich, sondern auch vorteilhaft und notwendig, um den harten Anforderungen an das Zahnsystem in der vorindustriellen Zeit standzuhalten. So können auch heutige Menschengruppen, wie etwa die australischen Ureinwohner oder ländliche Nigerianer, ihre Weisheitszähne noch gut nutzen.5 6
Dass heute bei vielen die Weisheitszähne keinen Nutzen erfüllen liegt nicht an der hypothetischen Makroevolution, sondern an unserer, für das Gebiss schonenden, „modernen“ Nahrung.
Ein ausführlicherer Artikel zum Thema ist hier zu finden.
4. Das Steißbein:
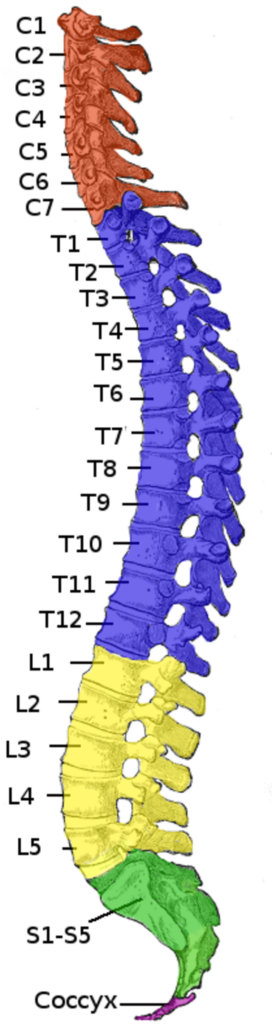
Das Steißbein wird als Überrest eines früheren Schwanzes interpretiert, da es sich ungefähr dort befindet, wo bei Affen mit Schwanz der Schwanz anfängt. Da es jedoch wichtige Funktionen erfüllt, ist dies nicht die einzig mögliche Interpretation:
Das Steißbein dient als Ansatzpunkt für verschiedene Bänder und Muskeln des Beckens, Beckenbodens und der Hüftgelenke. Die Bänder und Muskeln in diesem Bereich sind wichtig, um die Organe an ihrem Platz zu halten, da das Becken nach unten offen ist.
Weiter nimmt es die Kräfte der angewachsenen Muskeln, Bänder und Sehnen des Beckenbodens, des Beckens und der Hüften auf und ist somit wichtig für die Statik des Körpers und den aufrechten Gang.
Die gelenkige Verbindung zwischen dem Kreuzbein und den ersten beiden Wirbeln des weiblichen Steißbeins ist außerdem bei der Geburt wichtig:
Wenn der Kopf des Säuglings durch den Geburtskanal kommt, bewegt sich durch den Druck die Spitze des Steißbeins um ca. 2 Zentimeter nach hinten und vergrößert so den Beckenausgang, was den Durchtritt des Säuglings erleichtert. Diese Beweglichkeit verdankt das Steißbein seiner Herkunft aus der Wirbelsäule.
Auch das Steißbein ist ohne weiteres mit Schöpfung kompatibel. Die Interpretation als Rest eines Schwanzes ist rein subjektiv, basierend auf der vorausgesetzten Annahme, dass der Mensch von Affenartigen abstamme. Objektiv gesehen muss die Wirbelsäule halt einfach irgendwo enden.
Etwas ausführlicher zum Steißbein hier.
5. Die Abdominalknochen der Wale:
Abdominalknochen (Hüftknochen) bei Walen sind kleine Knochen ohne Verbindung zur Wirbelsäule im hinteren Teil des Wals. Sie werden als Überreste eines hypothetischen früheren Beckengürtels interpretiert:
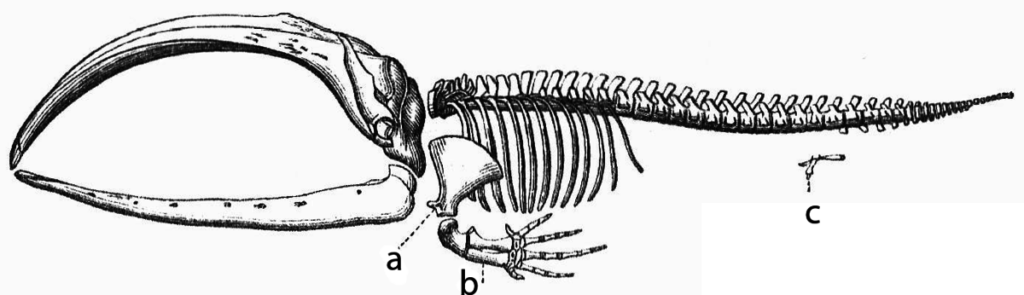
Doch auch die Funktionslosigkeit dieser Struktur konnte widerlegt werden. Die Abdominalknochen unterstützen die Muskelatur der Geschlechtsorgane, sind dementsprechend bei männlichen und weiblichen Walen unterschiedlich ausgebildet und spielen somit eine lebenswichtige Rolle bei der Paarung.7 In einer Studie von 2014 konnte außerdem ein Zusammenhang, zwischen Größe und Form der Abdominalknochen und dem Paarungsverhalten der Wale festgestellt werden.8
Somit ist es inzwischen sinnlos Abdominalknochen als Rudimente einzustufen. Nichts spricht dagegen, dass sie geschaffen wurden.
Etwas ausführlicher dazu hier.
6. Die menschliche Körperbehaarung:
Evolutionisten sehen in der spärlichen Körperbehaarung des Menschen einen funktionslosen Überrest eines früheren Fells. Zunächst hat der Mensch fast überall Haare. Neben den gut sichtbaren Haaren haben wir außer an den Handflächen und Fußsohlen überall winzige farblose Haare. Doch wie so oft machen Naturalisten es sich auch hier zu einfach. Ein wichtiger Unterschied zwischen uns Menschen und Tieren mit Fell ist die Art, wie wir uns abkühlen.
Die meisten Tiere können nicht schwitzen und brauchen dementsprechend ein dichtes Fell zur Wärmeisolierung. Sie kühlen sich dafür mit hecheln ab. Der Mensch jedoch kann schwitzen und wird durch das Verdampfen des Schweißes auf der Haut abgekühlt. Ein langes Fell wäre hierbei hinderlich.
Ein weiterer Unterschied ist, dass Tiere ihr Fell auch als Schutz vor der UV-Strahlung der Sonne brauchen, während wir Menschen uns hauptsächlich durch das Bräunen unserer Haut und mit Kleidung schützen. – nur unser Kopfhaar erfüllt ebenfalls diese Funktion.
Außerdem erfüllen unsere Haare – inklusive der winzigen, farblosen Haare – eine sensorische Funktion: Alle Haarfollikel (wo die Haare in der Haut verankert sind) sind mit Sinnesnerven ausgestattet und können mit Bewegungsmeldern verglichen werden. Werden die Haare durch irgendetwas bewegt, werden sensorische Signale an unser Gehirn gesendet. Das kann kaum als rudimentär bezeichnet werden.
Weiter sind die Haarfollikel wichtig bei der Wiederherstellung der Hautoberfläche nach Verletzungen. Sie dienen nämlich als wichtige Quelle der Epidermiszellen für die Wiederherstellung der Hautoberfläche. Ohne diese wäre selbst bei Schürfwunden eine Hauttransplantation notwendig.
An der Basis jeden Haares befinden sich Talgdrüsen, die ein öliges Sekret namens „Talg“ absondern. Dieser schützt die Haut vor Austrocknung und hält sie geschmeidig. Die Haare dienen dabei als „Leitbahnen“, über die der Talg auf die Hautoberfläche transportiert wird.
Die oft erwähnte Gänsehaut ist ebenfalls nicht funktionslos. Die Muskeltätigkeit der Hautmuskeln generiert eine gewisse Wärme und die dadurch aufgestellten Haare warnen die Person vor einem Körpertemperaturproblem.9 Weiter tritt sie auch bei emotionalen Reizen wie Freude oder Angst auf und kann als Warnhinweis dienen. Das kann kaum als Rest einer früheren Wärmefunktion gedeutet werden.
Hinzu kommt, dass Mann und Frau bekanntlich eine unterschiedlich starke Körperbehaarung besitzen. Aus Schöpfungsperspektive kann man das als eine vom Schöpfer gewollte optische Unterscheidung von Mann und Frau deuten.
Die menschliche Körperbehaarung ist keineswegs rudimentär und ohne weiteres mit einem Schöpfer vereinbar.
Weitere Informationen hier.
7. Darwins Ohrhöcker:
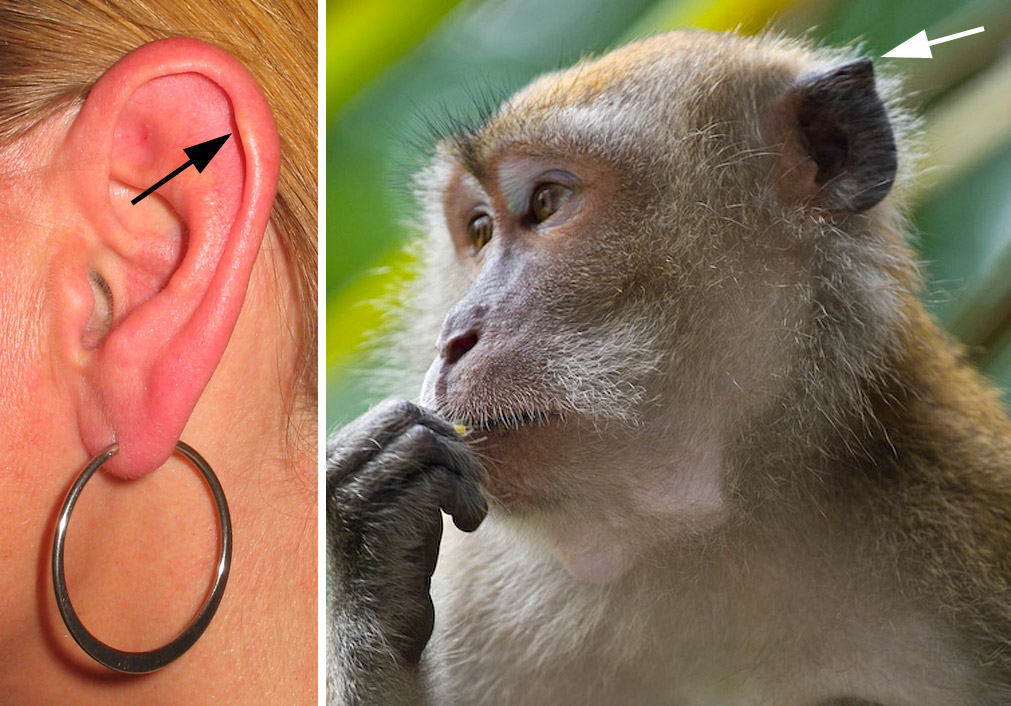
Viele Tiere können ihre Ohren bewegen. Wir können das zwar nicht, haben aber einen angeblich funktionslosen Höcker aus Knorpel am Rand unserer Ohrmuschel, der ein Überbleibsel dieser Fähigkeit sein soll.
Doch auch hier ist die Funktion inzwischen bekannt. Die neuronalen Schaltkreise der Ohrmuskeln in unserem Gehirn führen bei plötzlichen Geräuschen dazu, dass unsere Augen sich reflexartig zur Seite bewegen. Dieser Reflex geht mit einer leichten Krümmung der Ohrmuschel einher.10 Der Höcker trägt zusammen mit anderen Vertiefungen und Faltungen der Ohrmuschel zum räumlichen Hören bei.
Auch der Ohrhöcker ist kein eindeutiges Rudiment und völlig mit Schöpfung kompatibel.
8. Flugunfähige Vögel:
Einige gefiederte Vögel sind nicht in der Lage zu fliegen, da ihre Flügel nicht zum Fliegen geeignet sind:


Evolutionsvertreter sehen in diesen Flügeln die Reste früherer flugtauglicher Flügel.
Trotzdem erfüllen die Flügel immer noch Funktionen. Beim Kiwi regulieren sie bspw. das Gleichgewicht. Weiter können Flügel bei flugunfähigen Vögeln zum Wärmen bei kaltem Wetter und zum Kühlen bei warmen Wetter eingesetzt werden. Sie können bei Stürtzen den Brustkasten schützen, bei Balzritualen zum Einsatz kommen, Küken in Schutz nehmen und zur Verteidigung dienen. Emus laufen bspw. mit aufgerissenem Schnabel und flatternden Flügeln auf ihre Feinde zu.
In diesem Fall ist es durchaus möglich, dass die Flügel zum Teil degeneriert sind. Das ist aber auch mit dem Schöpfungsmodell vereinbar und kein Indiz für Makroevolution. Da sie aber auch einige Funktionen erfüllen, könnten sie auch von Anfang an so geschaffen worden sein. Was zutrifft muss von Einzelfall zu Einzelfall geklärt werden. Wo flugunfähige Vögel auf Inseln leben könnte das z.B ein Indiz für Degeneration sein, da Vögel auf Inseln oft weniger bis keine Feinde haben und somit auch gefahrlos am Boden bleiben können.
Dass solche Flügel rudimentär sind ist möglich, aber nicht zwingend notwendig und letztlich für die Frage „Schöpfung oder Evolution?“ irrelevant.
9. Hervorstehende Eckzähne:
Naturalisten nehmen an, dass unsere hervorstehenden Eckzähne im Lauf der Evolution kleiner geworden sind. Der Grund liegt in der Tatsache, dass unsere Eckzähne kürzer sind als die Eckzähne der Affen und die vorausgesetzte Annahme, dass wir von Affenartigen abstammen.
Offensichtlich handelt es sich hier um eine subjektive Deutung aus Sicht der ET. Objektiv betrachtet macht unser Eckzahn nicht den Eindruck rückentwickelt zu sein und steht in keinem Widerspruch zum Schöpfungsmodell. Außerdem widerspricht die Tatsache, dass die frühesten angeblichen tierischen Vorfahren des Menschen – die Australopithecinen – kleinere Eckzähne als der Mensch hatten, der evolutionstheoretischen Deutung.
Somit handelt es sich hier weder um einen Evolutionsbeleg, noch um ein Problem für einen Schöpfer.
10. Das Nickhaut-Rudiment:

Die Nickhaut ist bei vielen Wirbeltieren ein durchsichtiges drittes Augenlied am nasenseitigen Augenwinkel, welches bei Bedarf zum Schutz über das ganze Auge geklappt werden kann. Der Mensch hat an den nasenseitigen Augenwinkeln ebenfalls eine Art Nickhaut, die bei uns aber nicht beweglich ist und von Evolutionsvertretern als der Rest einer früheren beweglichen Nickhaut interpretiert wird.

Doch auch hier handelt es sich nur um eine Deutungsmöglichkeit, denn unsere Nickhaut erfüllt den Zweck Fremdstoffe, die ins Auge gelangt sind, zu einer klebrigen Masse am Augenwinkel zu sammeln, wo diese dann gefahrlos entfernt werden können. Außerdem unterscheidet die menschliche Nickhaut sich strukturell deutlich von der Nickhaut der Vögel.
Die Deutung als Rudiment mag aus evolutionstheoretischer Sicht unter der Annahme eines teilweisen Funktionsverlustes oder Funktionswandels möglich sein, aber keine zwingende Notwendigkeit. Da die Nickhaut eine Funktion erfüllt, kann sie auch von Anfang an so geschaffen worden sein.
11. Augenreste bei Höhlenfischen:

Fische, die in Höhlen leben, sind blind und verfügen nur noch über Augenreste. Der Grund liegt darin, dass Fische in Höhlen wegen der Dunkelheit keine Augen brauchen, eine Rückentwicklung somit keinen Nachteil bringt und infolgedessen auch nicht ausselektiert wird. Im Gegenteil hat sie sogar einen Vorteil, da blinde Fische sich nicht die Augen verletzten können, wenn sie z.B. gegen Höhlenwände stoßen.
Doch ist auch diese Rückentwicklung mit dem Schöpfungsmodell vereinbar und kein Beleg für Makroevolution, da die Rückentwicklung nur innerhalb der entsprechenden Fischart stattgefunden hat und der im Schöpfungsmodell erwarteten Entwicklungsrichtung entspricht.
Mit einem Experiment wurde außerdem nachgewiesen, dass es diese Rückentwicklung nicht seit millionen Jahren geben kann:
Richard Borowsky und seine Kollegen gingen berechtigter Weise davon aus, dass bei verschiedenen Populationen verschiedene Mutationen zum Sehverlust geführt hatten. „Populationen“ meint hier verschiedene Gruppen der selben Art (in diesem Fall Astyanax mexicanus), die räumlich und / oder zeitlich voneinander getrennt sind.
Sie nahmen vier verschiedene Populationen des mexikanischen Höhlenfisches und kreuzten diese miteinander. Im Ergebnis konnten fast 40% der Nachkommen sehen.11 Da der Verlust der Sehfähigkeit bei den verschiedenen Populationen auf verschiedene geschädigte Gene zurück ging, konnten die Schäden des einen Elternteils jeweils durch die gesunden Gene des anderen Elternteils beim Nachkommen aufgehoben werden.
Dr. Richard Borowsky sagt: „Es werden nicht nur die Strukturen des Auges so weit wiederhergestellt, dass sie wieder funktionieren, sondern auch alle Verbindungen zum Gehirn, die für die ordnungsgemäße Verarbeitung von Informationen sorgen, […].12
Die Tatsache, dass Nachkommen mit funktionierenden Augen gezeugt werden konnten, lässt 2 Schlussfolgerungen zu:
- Die Erbinformation für funktionierende Augen ist nach wie vor im Erbgut der blinden Fische enthalten.
- Folglich kann diese Rückentwicklung noch nicht so alt sein. Bei einer Zeitspanne von millionen Jahren wäre dieser Bauplan längst ausselektiert worden. D.h. er wäre durch Mutationen weiter verkommen und da er nicht gebraucht wird, hätte die Selektion ihn nicht in der Population erhalten.
Blinde Höhlenfische sind also vollständig mit dem Schöpfungsmodell vereinbar. Etwas ausführlicher dazu in diesem Artikel.
12. Die Afterklauen der Python und Boa:
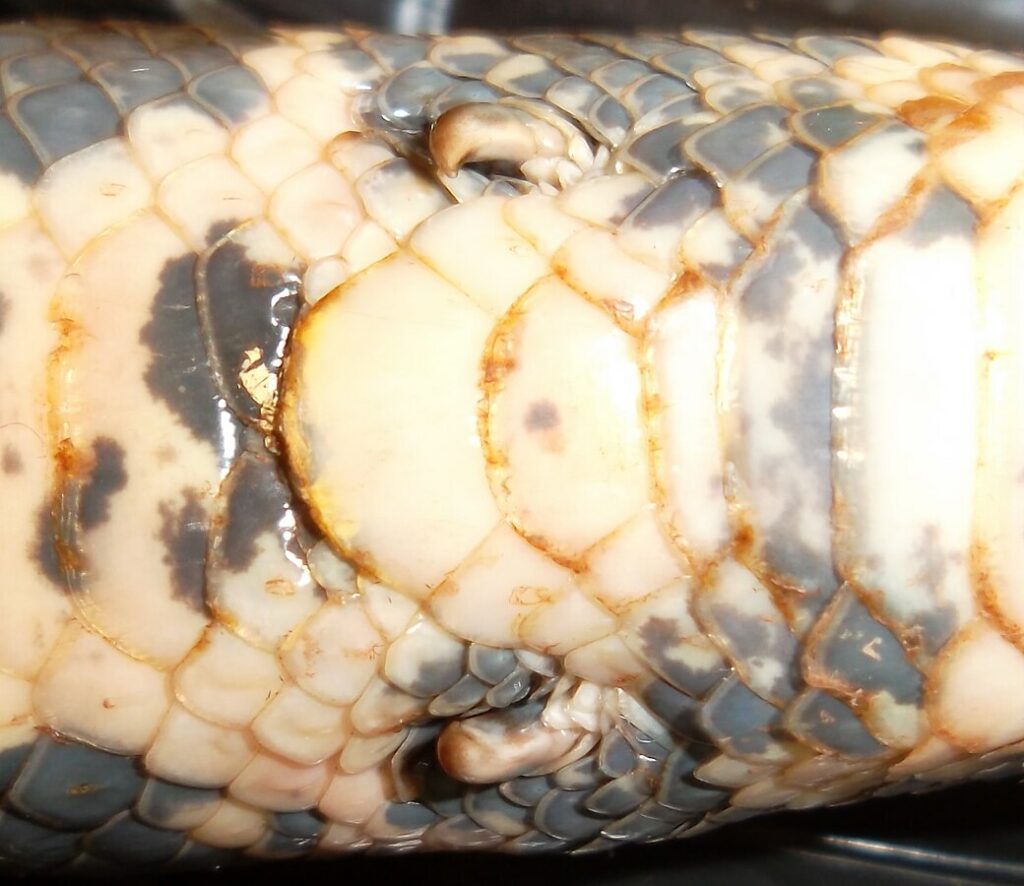
Pythons und Boas haben Afterklauen, die von Naturalisten als Reste früherer Beine interpretiert werden. Unabhängig davon, ob das stimmt, sind die Afterklauen beim Bewegen in Geäst und bei der Paarung hilfreich. Eine Interpretation als Rudiment ist also nicht zwingend.
Doch auch aus Sicht der biblischen Schöpfungslehre ist zu erwarten, dass Schlangen früher Beine hatten:
1.Mose 3,14 Da sprach Gott der HERR zur Schlange: Weil du dies getan hast, so sollst du verflucht sein mehr als alles Vieh und mehr als alle Tiere des Feldes! Auf deinem Bauch sollst du kriechen und Staub sollst du fressen dein Leben lang!
Die Bibel impliziert also, dass die Schlange zu Beginn mit Beinen geschaffen wurde. Sollte es sich also tatsächlich um ein Rudiment früherer Beine handeln, ist das sowohl im Rahmen des Evolutionsmodells, als auch im Rahmen des Schöpfungsmodells interpretierbar. Wobei anzumerken ist, dass eine Interpretation im Evolutionsmodell kein Beleg für Makroevolution ist, da etwas verloren ging und nicht neu entstanden ist.
Etwas ausführlicher dazu in diesem Artikel.
13. Blinde Maulwürfe:

Maulwürfe sind bekanntlich oft blind. Sie haben zwar Augen, diese sind aber von einer äußeren Hautschicht bedeckt und eignen sich nur zum Unterscheiden von Tag und Nacht.
Naturalisten sehen in diesen Augen ein Rudiment und manchmal werden sie auch als Konstruktionsfehler gegen intelligentes Design vorgebracht.
Zunächst wäre der Verlust der Sehfähigkeit, wie auch schon bei den Höhlenfischen erklärt, kein Problem für Schöpfung. Doch müssen die Augen des Maulwurfs überhaupt rudimentär sein?
Maulwürfe leben die meiste Zeit unter der Erde im Dunkeln, „normale“ Augen wären hier überflüssig. Zudem graben sie hauptsächlich mit dem Maul, hier wären normale Augen ein Risiko, da sie beim Graben verletzt werden und sich entzünden könnten.
Maulwürfe kommen zur Nahrungssuche, zur Fortbewegung und zur Paarung an die Oberfläche. Mit sehenden Augen müssten sie sich erst an das Licht gewöhnen und wären desorientiert, was für Tiere am unteren Ende der Nahrungskette durchaus lebensgefährlich sein kann.
So aber kommen blinde Maulwürfe bei Tageslicht besser zurecht, als sehende nachtaktive Nagetiere,13 was bedeutet, dass sie viel weniger Zeit zur Gewöhnung an das Tageslicht brauchen.
Neben der Unterscheidung von Tag und Nacht haben Augen bei Säugetieren außerdem eine Aufgabe als wichtiges Bindeglied zur Wärmeregulation, Fortpflanzung und zum Winterschlaf, die alle Informationen zur Veränderung der Tageszeit brauchen. Dementsprechend haben Maulwürfe funktionsfähige Netzhäute, die Melatonin pruduzieren, welches ebenfalls zur Wärmeregulierung beiträgt. So zeigte sich auch in einer Studie von 1990, dass die Entfernung der Augen zu einer Verschlechterung der Wärmeregulation führt.14
Wir sehen also, dass die Augen des Maulwurfs optimal für den Lebensraum des Maulwurfs konzipiert und keineswegs ein Konstruktionsfehler sind. Egal ob der Maulwurf blind geschaffen wurde oder seine Sehfähigkeit durch Degeneration verloren hat, beides es mit dem Schöpfungsmodell vereinbar.
Ausführlicher wird die Thematik in diesem Artikel behandelt.
14. Der Greifreflex:

Neugeborene Kinder besitzen einen Greifreflex, wobei das Kind automatisch mit der Hand zugreift, wenn etwas seine Handinnenfläche berührt. Dieser Reflex läuft unbewusst ab und kann bis zum 9. Monat nach der Geburt auftreten. Evolutionsvertreter sehen den Ursprung dieses Reflexes in dem typischen greifen von Ästen bei Affen und dem Festkrallen junger Affen am Fell der Mutter.
Doch entwicklungsphysiologische Zusammenhänge lassen an dieser Erklärung Zweifel aufkommen. Während der Frühdifferenzierung führen die Armanlagen eine Entwicklungsbewegung durch, welche „Wachstumsgreifen“ genannt wird.
Auf unbewusster embryonaler Ebene im Rahmen der Armentwicklung erfolgt dadurch eine Vorbereitung auf das spätere bewusste Greifen.
Ab der 5. Schwangerschaftswoche beginnt das Wachstum des Ärmchens des Embryos zur Seite und anschließend nach vorn. Danach wird der Ellenbogen leicht gekrümmt, wodurch eine leichte Annäherung der Hand an die vordere Brustwand stattfindet. Anschließend wird die Hand zum Mund geführt.
Hier ist nun ab der 7. Schwangerschaftswoche eine reifungsbedingte und wachstumsstimulierende Greifbewegung beobachtbar. Diese Greifbewegung ist beim Neugeborenen noch als Handgreifreflex auslösbar und verschwindet mit der Zeit durch die weitere Reifung des Gehirns, des Nerven- und Bewegungssystems. Der Fußgreifreflex wird übrigens durch ähnliche Zusammenhänge erklärt.
Diese funktionelle Erklärung sowie die Tatsachen der ontogenetischen Reifungsprozesse machen die Deutung als Rudiment überflüssig. (Ontogenese = Entwicklung eines einzelnen mehrzelligen Organismus) Der Greifreflex steht in keinerlei widerspruch zu Schöpfung.
Etwas ausführlicher zu diesem Beispiel in diesem Artikel.
15. Die männlichen Brustwarzen:
Männliche Brustwarzen sind ebenfalls ein beliebtes Rudiment von Evolutionsvertretern. Da sie offensichtlich keine Milch geben, werden sie als nutzlos angesehen. Die Annahme liegt zwar auf den ersten Blick nahe, aber stimmt das wirklich?
Mann und Frau sind in den ersten Stadien der Embryonalentwicklung physiologisch identisch. Brustwarzen gehören zum Grundbauplan des Menschen und sind bereits vorhanden, wenn die Geschlechtsentwicklung beginnt.
Sowohl die männlichen als auch die weiblichen Brustwarzen enthalten reichlich Nervengewebe und sind deshalb sehr berührungsempfindlich.15 Sie werden von mehreren Nerven innerviert,16 was für sich schon auf eine Funktion schließen lässt. Die Brustwarzen des Mannes haben mehrere Funktionen, sind aber vor allem an der sexuellen Stimulation beteiligt17,18 und ebenso empfindlich wie weibliche Brustwarzen.19 Die Brustwarzen der Frau haben aber mehr und größere erogene Zonen als die des Mannes.
Die Bedeutsamkeit der männlichen Brustwarzen wird auch durch die Versuche angezeigt, den Brustwarzen-Areolarkomplex nach einem Unfall oder einer Erkrankung wiederherzustellen.20 Nur beim Menschen sind Brüste und Brustwarzen am Geschlechtsverkehr beteiligt, es gibt keine Hinweise auf eine Evolution dieser Funktion bei niederen Primaten.
Interesannt wäre außerdem von was genau männliche Brustwarzen ein Rudiment sein sollen, worauf gewöhnlich aber nicht eingegangen wird.
Männliche Brustwarzen sind also durchaus funktionell und keineswegs als rudimentär einzustufen.
Ein ausführlicherer Artikel zu diesem Thema hier.
16. Schulter- und Beckenknochenreste bei Blindschleichen:

Blindschleichen sehen zwar auf den ersten Blick wie Schlangen aus, werden aber zu den Echsen gerechnet. Merkmale wie bewegliche Augenlieder und Ohröffnungen stellen sie eher zu Echsen, als zu Schlangen, denen diese Merkmale fehlen. Außerdem sind ihre Kiefer weniger elastisch und ihre Fortbewegung weniger fließend und steifer als bei Schlangen.
Blindschleichen besitzen auch Knochen, die als funktionslose Reste von Schulter und Becken interpretiert werden. Erklärt werden diese (mutmaßlichen) Rudimente mit einer Abstammung von Echsen. Zunächst kann das Vorhandensein einer Funktion methodisch nicht definitiv ausgeschlossen werden. Läge doch eine Funktion vor, könnten Blindschleichen auch so geschaffen worden sein.
Sollten die Knochen jedoch tatsächlich Reste früherer Beine sein, könnte man aus schöpfungswissenschaftlicher Sicht diskutieren, ob mikroevolutive genetische Variationsmechanismen (Rekombination; Epigenetik: springende Gene,…; Variationsprogramme), die Merkmale, die evolutionstheoretisch als Neuerwerb während der Rudimentation interpretiert werden, auch innerhalb eines Grundtyps hervorbringen könnten. Wie wir heute wissen, ist auf mikroevolutiver Ebene (ohne Informationszunahme) ein beachtliches Maß an Variation und latenten Programmen möglich.
Im konkreten Fall der Blindschleiche müsste dieser Ansatz aber noch genauer anhand aktueller Daten geprüft werden.
Zudem ist eine Abstammung von (anderen) Echsen aus Schöpfungssicht nicht zwingend. Wenn Blindschleichen als eigener Grundtyp verstanden werden, der ggf. früher laufen konnte, könnte dieser morphologisch (körperbaulich) heutigen Blindschleichen ähnlicher gesehen haben als andere Echsen heutigen Blindschleichen und die mikroevolutiv zu erklärenden Unterschiede wären geringer. Die Ähnlichkeiten zwischen Echsen und Blindschleichen wären dann geschaffene Konvergenzen.
Blindschleichen sind also im Rahmen beider Modelle interpretierbar.
Weitere (angebliche) Rudimente werden mit der Zeit ergänzt. (Zuletzt aktualisiert am 03.04.2025)
Allgemeines:
Rudimente werden als eine der besten Belege für Evolution angeführt. Doch nüchtern betrachtet sind sie das nicht. Wo immer ein echtes Rudiment vorliegt, zeugt es von Degeneration, nicht von Evolution. Die ET braucht aber Weiterentwicklung, nicht Rückentwicklung. Rückentwicklung ist die im Schöpfungsmodell erwartete Entwicklungsrichtung und dementsprechend sind hier auch Rudimente denkbar. Rudimente könnten in Einzelfällen nur dann (eindeutig) auf Makroevolution hindeuten, wenn die Rückentwicklung über Grundtypgrenzen hinweg stattgefunden hätte – vielleicht noch in Verbindung mit der Entstehung neuer Organe oder Strukturen -, wie es etwa bei den Hüftknochen der Wale der Fall wäre.
Doch auch wenn es solche scheinbar eindeutigen Rudimente geben sollte, haben Evolutionsvertreter das Problem, dass sie die Funktionslosigkeit eines Organs nicht empirisch (mit Beobachtungen und Experimenten) nachweisen können. Die Funktionslosigkeit ist lediglich eine subjektive Annahme unter Voraussetzung der ET. Faktisch könnte auch einfach eine bislang unbekannte Funktion vorliegen, wie es schon bei so vielen angeblichen Rudimenten der Fall war.
In dem Punkt ist der Naturalismus auch ein Hindernis für die natuwissenschaftliche Forschung und den Erkenntnisgewinn. Denn warum sollte man etwas untersuchen, von dem man überzeugt ist, es sei ein nutzloses Rudiment? Hier spornt vielmehr Wissenschaft auf Grundlage der Schöpfungslehre zu weiterer Forschung an.
Letzlich sind eventuelle echte Rudimente also Deutungssache und eher schwache Argumente für Makroevolution, als starke.
Fußnoten:
- Paul Rea, “Vagus Nerve,” Essential Clinical Anatomy of the Nervous System (Massachusetts: Academic Press, 2015), 10.7.1. ↩︎
- Yong Tae Hong und Ki Hwan Hong, “The Relationship Between the Non-recurrent Laryngeal Nerve and the Inferior Thyroid Artery,” Indian Journal of Surgery 80, no. 2 (April 2018): 109–112. ↩︎
- Cramon-Taubadel, “Global Human Mandibular Variation Reflects Differences in Agricultural and Hunter-Gatherer Subsistence Strategies” PNAS 108, no. 49 (December 2011): 19546–19551, doi:10.1073/pnas.1113050108; Anita Sengupta, David K. Whittaker, Geraldine Barber, Juliet Rogers, und Jonathan H. Musgrave, “The Effects of Dental Wear on Third Molar Eruption and on the Curve of Spee in Human Archaeological Dentitions,” Archives of Oral Biology 44, no. 11 (1999): 925–934, doi:10.1016/S0003-9969(99)00095-3. ↩︎
- Cramon-Taubadel, “Global Human Mandibular Variation Reflects Differences in Agricultural and Hunter-Gatherer Subsistence Strategies”; and Y. Kaifu, K. Kasai, G. C. Townsend, and L. C. Richards, “Tooth Wear and the ‘Design’ of the Human Dentition: A Perspective from Evolutionary Medicine,” American Journal of Physical Anthropology, 122 (2003): 47–61, doi:10.1002/ajpa.10329. ↩︎
- R. S. Corruccini, “Australian Aboriginal Tooth Succession, Interproximal Attrition, and Begg’s Theory,” American Journal of Orthodontics and Dentofacial Orthopedics 97, no. 4 (1990): 349–357, doi:10.1016/0889-5406(90)70107-N. ↩︎
- S. A. Odusanya und I. O. Abayomi, “Third Molar Eruption Among Rural Nigerians,” Oral Surgery, Oral Medicine, and Oral Pathology 71 (1991): 151–154, doi:10.1016/0030-4220(91)90457-N. ↩︎
- Siehe Wieland, C., 1988. The Strange Tale of the Leg on a Whale. Creation
20(3):10-13. ↩︎ - Dines JP, Otárola-Castillo E, Ralph P, Alas J, Daley T, Smith AD & Dean MD (2014) Sexual selection targets cetacean pelvic bones. Evolution,doi: 10.1111/evo.12516 ↩︎
- Bergman J & Rowe G (1990) „Vestigal organs“ are fully functional. Creation Reserch Society Monograph Series No. 4. Terre Haute, IN. ↩︎
- Steven Hackley, “Evidence for a Vestigial Pinna-Orienting System in Humans,” Psychophysiology 52, no. 10 (2015): 1263–1270, doi:10.1111/psyp.12501. ↩︎
- New York University, Progeny of blind fish can regain their sight, ScienceDaily Jan 8, 2008 http://www.sciencedaily.com/releases/2008/01/
080107120911.htm Die Arbeit wurde offiziell in einer Ausgabe von Current Biology veröffentlicht. ↩︎ - Originalzitat: „Not only are the structures of the eye restored to the point where they regain function, but all the connections to the brain for proper processing of information not used for that enormous length of time are restored.“; Gleiche Quelle wie in Fußnote 10. ↩︎
- Reut Avni, Yael Tzvaigrach und David Eilam, “Exploration and navigation in the blind mole rat (Spalax ehrenbergi): global calibration as a primer of spatial representation,” Journal of Experimental Biology 211, (2008): 2825, https://jeb.biologists.org/content/jexbio/211/17/2817.full.pdf. ↩︎
- Satrajit Sanyal, HG Jansen, Willem J. Degrip, und Eviatar Nevo, “The Eye of the Blind Mole Rat, Spalax ehrenbergi, Rudiment With Hidden Function?” Investigative Ophthalmology & Visual Science 31, no. 7, (July 1990): 1402-1403, https://www.researchgate.net/publication/21014181_The_eye_of_the_blind_mole_rat_Spalax_ehrenbergi_Rudiment_with_hidden_function. ↩︎
- Sarhadi, N.S. and Lee, F.D., An anatomical study of the nerve supply of the breast, including the nipple and areola, British J. Plastic Surgery 49(3):156–164, 1996.
Sarhadi, N.S., Shaw-Dunn, J. and Soutar, D.S., Nerve supply of the breast with special reference to the nipple and areola: Sir Astley Cooper revisited, Clinical Anatomy 10(4):283–288, 1997.
Sykes, P.A., The nerve supply of the human nipple, J. Anatomy, 105(1):201, 1969.
Wuringer, E., Mader, N., Porsch, E. and Holle, J., Nerve and vessel supplying ligamentous suspension of the mammary gland, Plastic and Reconstructive Surgery 101(6):1486–1493, 1998.
Robinson, J.E. and Short, R.V., Changes in breast sensitivity at puberty, during the menstrual cycle, and at parturition, British Medical Journal 1(6070):1188–1191, 1977.
Kapdi, C.C. and Parekh, N.J., The male breast, Radiology Clinical North America 21(1):137–148, 1983. ↩︎ - Sykes, P.A., The nerve supply of the human nipple, J. Anatomy, 105(1):201, 1969. ↩︎
- Sloand, E., Pediatric and adolescent breast health, Lippincotts Primary Care Practice 2(2):170–175, 1998. ↩︎
- Masters, W. and Johnson, V., Human Sexual Response, Little Brown, Boston, 1966. ↩︎
- Stoppard, M., The Breast Book: The Essential Guide to Breast Care & Breast Health for Women of All Ages, Dorling Kindersley, New York, 1996. ↩︎
- DeBono, R. and Rao, G.S., A simple technique for correction of male nipple hypertrophy: the ‘sinusoidal’ nipple reduction, Plastic Reconstruction Surgery 100(7):1890–1892, 1997.
Aiache, A.E., Male chest correction. Pectoral implants and gynecomastia, Clinical Plastic Surgery 18(4):823–828, 1991.
Vasconez, H.C. and Holley, D.T., Use of the tram and latissimus dorsi flaps in autogenous breast reconstruction, Clinical Plastic Surgery 22(1):153–166, 1995.
Kincaid, S.B., Breast reconstruction: a review, Annuals of Plastic Surgery 12(5):431–448, 1984.
Liebau, J., Machens, H.G. and Berger, A., Gynecomastia of the male nipple, Annuals of Plastic Surgery 40(6):678–681, 1998. ↩︎