Die Entstehung der Feder
Dieser Artikel befasst sich mit der Entstehung der flugfähigen Feder und basiert auf dem Buch „Schöpfung ohne Schöpfer? – Eine Verteidigung des Design-Arguments in der Biologie“ von Dr. Reinhard Junker und Dr. Markus Widenmeyer.
Die gängige Auffassung lautet, dass Federn evolutiv, beginnend mit vogelähnlichen Dinosauriern, entstanden seien. Das gründet sich jedoch in erster Linie auf der prinzipiellen Grundannahme, dass alles durch rein natürliche Prozesse entstanden sei.
Um die Herkunft der Feder erklären zu können, muss diese zunächst genau untersucht werden: Welche grundsätzlichen Voraussetzungen müssen flugtaugliche Federn erfüllen? Wie müssen sie im Körper verankert sein? Wie funktioniert ihre Steuerung und wie beweglich müssen sie sein? Usw.
Eine Feder besteht äußerlich aus folgenden Bestandteilen:
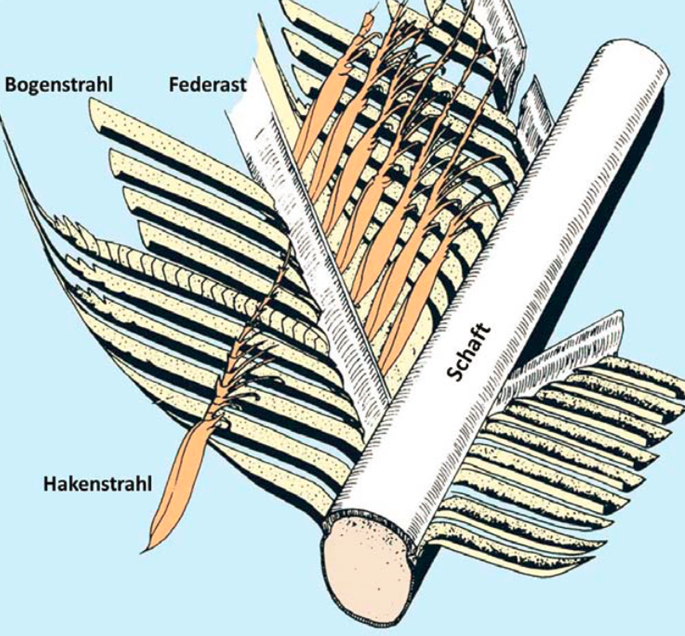
Es gibt keine komplexere Körperbedeckung als das Federkleid eines Vogels. Zunächst muss die Feder aus dem richtigen Material bestehen. Über 80% des Federmaterials bestehen aus langen Fasern des speziellen Proteins beta-Keratin. Beta-Keratin ist sowohl in Federschaft, und -ästen, als auch in den Federstrahlen verbaut. Nur so können die Federn ihre besonderen Eigenschaften aufweisen: robust, biegsam, leicht, knickfest, drehbar.
Die Fasern sind hierarchisch in 3 Stufen gebündelt, stehen zum Teil über Kreuz und einzelne Faserzüge zweigen vom Schaft in die Äste ab. Die Federäste wurzeln also im Schaft, was zur Knickfestigkeit und Stabilität beiträgt.
Die Federstrahlen sind ganz besonders gebaut: Sie gehen von den Ästen nach beiden Seiten ab. Auf der einen Seite befinden sich Hakenstrahlen mit winzige Häkchen, welche mit den Bogenstrahlen des benachbarten Astes verhaken – wie bei einem Reisverschluss. Dank dieses dichten Reisverschlusses ist die Federfahne praktisch luftundurchlässig. Wird die Feder zu stark belastet, reißen die Reisverschlüsse kontrolliert auf und verhindern so irreparable Schäden. Der Reisverschluss kann vom Vogel mithilfe seines Schnabels einfach wieder geschlossen werden.
Wie der Rest der Feder ist auch der Schaft besonders gebaut: Innen ist er mit einem Netzwerk aus schaumähnlichen Fasergeflecht gefüllt. Diese sind mit einem chemischen Stoff beschichtet, durch den der Federschaft unter schwachem Druck steht. Das hat zur Folge, dass er nicht so leicht geknickt werden kann und bei Verbiegung wieder in die ursprüngliche Form zurückgeht.
Obwohl sich diese Anforderungen nur auf den von außen sichtbaren Teil der Feder beziehen, sind diese schon komplex genug, um eine evolutive Entstehung in Zweifel zu ziehen bzw. um einen Hersteller in Betracht zu ziehen. Denn feinabgestimmte Komplexität zeugt objektiv betrachtet von einem Hersteller – wie auch die feinabgestimmte Komplexität einer Uhr von einem Uhrmacher zeugt.
Interessant sind aber auch die übrigen Bestandteile einer flugfähigen Feder:
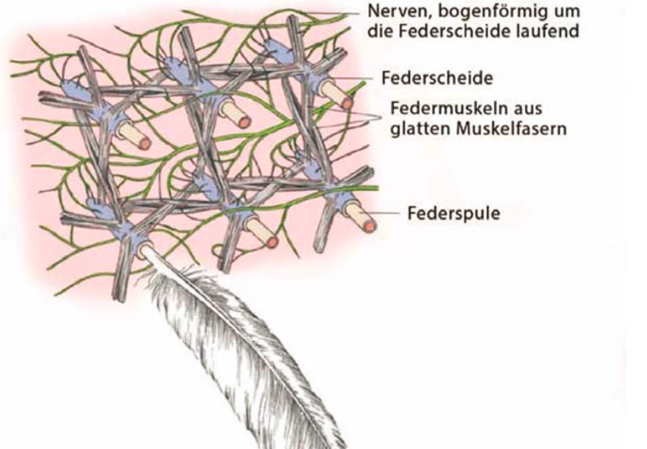
Der Follikel ist die Einsenkung in den Körper, in welche die Feder eingebettet ist. Er ist entscheidend für die Funktionalität der Federn, wie auch bei der ontogenetischen Entwicklung (Individualentwicklung) und der Mauser. Der Follikel verankert außerdem die Feder optimal im Körper – nicht zu locker und nicht zu fest. Dafür zuständig sind Bindegewebe und Fettpolster. Ein Muskelgeflecht an der Federspule (eingesenkter Teil der Feder) ermöglicht das koordinierte Bewegen der Federn. Die Muskeln wiederum benötigen Blutgefäße zur Versorgung und Nervenbahnen zur Reizweiterleitung, um zu funktionieren.
Sinneskörperchen und spezielle Fadenfedern registrieren die Positionen der einzelnen Federn. Diese Information wird weitergeleitet und in Sekundenbruchteilen verarbeitet, damit die gerade passenden Federbewegungen ausgeführt werden können.
Aber nicht nur der Aufbau der Feder an sich ist entscheidend, sondern auch die Pflege derselben. Denn was nützt ein anfangs perfektes Federkleid, wenn es durch Abnutzung und Beschädigung unbrauchbar wird und nicht erneuert wird?
Hier kommt die Mauser ins Spiel: Das abgenutzte Federkleid wird in der Mauser gegen ein neues ausgetauscht – was ebenfalls ein komplizierter Vorgang ist. Zudem produzieren Vögel in der Bürzeldrüse ein öliges Gemisch, welches sie mit dem Schnabel aufnehmen, um damit das Gefieder zu pflegen und zu imprägnieren.
Was natürlich auch noch gebraucht wird, ist eine Bauanleitung für den Aufbau der Federn während der individuellen Entwicklung. – was ebenfalls ein sehr anspruchsvoller und komplizierter Vorgang ist. Doch auch der (Fein)Bau und die Steuerung der einzelnen Federn ist wieder nur ein Teil der Notwendigkeiten für die Flugfähigkeit:
Denn auch das Federkleid muss insgesamt passend ausgebildet sein. Dies benötigt ebenfalls eine anspruchsvolle Steuerung, bei welcher viele Muskelpartien und die einzelnen Teile des Federkleids zusammenwirken müssen. Das macht wiederum zahlreiche Regelkreise und Rückkopplungsmechanismen erforderlich.
Zudem muss der ganze Vogel in Leichtbauweise „ausgeführt“ sein. So muss z.B. das ganze Skelett leicht, sehr robust und relativ starr konstruiert sein.
Aus den oben genannten Gründen weisen viele Wissenschaftler auf eine „Synorganisation“ hin. Das bedeutet, dass die einzelnen Module und Ebenen (Baumaterial bis Verhalten) in Abhängigkeit voneinander funktionieren. Sie können weder isoliert voneinander verstanden werden noch isoliert voneinander entstanden sein. Denn mit nur einem Teil dieser Voraussetzungen wäre der Vogel nicht (dauerhaft – siehe Mauser) flugfähig – was ja das Ziel ist. Zur hypothetischen evolutiven Entstehung später mehr.
Insgesamt stellt das Federkleid also eine nichtreduzierbare Gesamtorganisation dar, was ein Indiz für einen Hersteller darstellt.
Bevor wir nun zu der naturalistischen Entstehungshypothese kommen, sollen Kriterien genannt werden, an denen diese geprüft werden kann:
Die Triebfeder von Makro-Evolution sollen vorteilhafte Mutationen (99% der Mutationen sind schädlich) und Selektion sein. Vorteilhafte Mutationen verschaffen dem Lebewesen einen Überlebensvorteil, durch den sich solche Mutationen gegenüber anderen durchsetzen können und über Generationen in der Population fixiert werden, da Artgenossen ohne diese Mutation mit der Zeit aussterben (Selektion).
Mutationen treten ohne Zielorientierung auf, d.h. es existiert kein Plan, für welche Konstruktion sie später gebraucht werden könnten. Daraus folgt, dass i.d.R. jede fixierte Mutation dem Lebewesen sofort einen , Nutzen bringen muss, da sie sonst ausselektiert würde oder zufällig verloren gehen könnte (bei neutralen Mutationen).
Die von der Mehrheit bevorzugte hypothetische evolutionäre Entstehung ist ein Modell nach Prum:
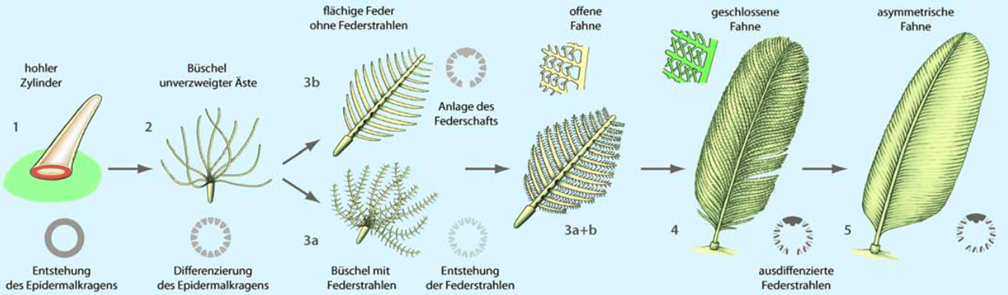
- Es bildet sich eine in die Haut einsinkende enge Röhre. Die Einsenkung soll später zum Follikel werden. Zudem entsteht an der Basis der Epidermalkragen.
- Daraus folgt eine daunenartige büschelige Feder ohne Verzweigungen. Zudem differenziert sich der Epidermalkragen in 2 Schichten.
- Entweder bildet sich zuerst der Federschaft mit Seitenästen in flächenförmiger Anordnung oder es bilden sich zuerst an den Daunenfedern Abzweigungen (zukünftige Federstrahlen). Beide Varianten zusammen ergäben dann die erste Deckfeder mit noch unverbundenen Federstrahlen.
- Es kommt zur Differenzierung von Bogen-und Hakenstrahlen. Die Konturfeder ist in diesem Stadium noch symmetrisch und somit nicht flugtauglich.
- Zum Schluss kommt eine Asymmetrie der Feder dazu, womit sie flugtauglich wird.
Dieses Modell ist jedoch aus folgenden Gründen nicht realistisch:
- 5 Stadien sind viel zu grob, um eine evolutionäre Entstehung realistisch modellieren zu können. Es werden weder ein Großteil der anatomischen Verhältnisse beachtet, noch die detaillierten histologischen Verhältnisse (Gewebe im Bereich des Follikel) oder die Synorganisation der Feder, usw. Z.B. muss die Steuerung der Federbewegung – welche an sich schon sehr komplex ist – und ihre Regulation auf die Koordination des Flugapparates abgestimmt sein. Auch andere Modelle mit bis zu 8 Stadien sind viel zu grob.
- Die erforderlichen Änderungen von Stufe zu Stufe sind enorm. Am Bsp. des ersten hypothetischen Stadiums: In diesem Modell nimmt die Entstehung des Follikels eine Schlüsselrolle ein und macht einen großen Sprung erforderlich. Große Sprünge durch Mutation sind jedoch nicht experimentell belegt, sie sind reine Spekulation. Zusätzlich ergibt die Einsenkung nur Sinn, wenn die feine Röhre darin bewegt werden kann. Sonst würde ein einfacher Auswuchs reichen. Für die Einsenkung hingegen werden Muskeln, Sehnen, Innervierung, Sinnesorgane usw. benötigt – zwar nicht unbedingt im heutigen Ausmaß, doch ohne ein Mindestmaß an Vernetzung und Abstimmung kann keine Funktionalität gewährleistet werden und ein selektiver Vorteil würde somit unmöglich. Es werden wichtige Teilaspekte ignoriert, weshalb das Modell das Erklärungsziel (Flugtauglichkeit der Feder) verfehlt.
- Zum Teil wird mit den einzelnen Schritten eine Zielorientierung impliziert, welche es im naturalistischen Denkrahmen jedoch nicht geben darf. Das kann als Hinweis verstanden werden, dass Naturalisten eigentlich keine Ahnung haben, wie Neuheiten auf natürlichem Wege entstehen sollen. Wieder am Bsp. des ersten Stadiums: Warum sollten eine Einsenkung und eine hohle Röhre in diesem Stadium einen selektiven Vorteil haben? – Zielorientierung gibt es ja nicht. Warum keine ganz gewöhnliche kompakte Röhre? Die hohle Röhre ist im ersten Stadium des Modells enthalten, da sie später notwendig für die Flugtauglichkeit ist. Ein zukünftiger Nutzen darf im Naturalismus aber keine Erklärung sein! Selektion plant nicht, sie wählt nur aus, was in der jeweiligen aktuellen Situation am vorteilhaftesten ist.
Nun noch kurz zum Fossilbericht: Es gibt zwar Dinosaurierfossilien mit Körperbedeckungen, die den einzelnen Stadien entsprechen könnten, doch passen die Abfolgen nicht zum Modell. So hat schon der Archaeopteryx (frühe fossile Form mit Federn) asymmetrische, möglicherweise flugtaugliche Federn, während fast alle Dinosauriergattungen mit einfacherer Körperbedeckung in jüngeren Kreideschichten gefunden wurden. Zudem ist damit nicht beantwortet, wie der evolutionäre Umbau funktionieren soll. Das Vorkommen von angeblichen Zwischenformen (dazu noch in verkehrter Reihenfolge) ist noch kein Beleg für einen ausreichend wahrscheinlichen Mechanismus. Somit ist eine rein natürliche und zukunftsblinde Entstehung von flugtauglichen Federn nach wie vor im Wesentlichen ungeklärt.
Fazit:
Die flugtaugliche Feder ist ein Meisterwerk an Vernetzung, (Fein)Abstimmung und Komplexität. Eine rein natürliche Entstehung konnte bislang nicht plausibel begründet werden. Wenn man aber die Grenzen des naturalistischen Denkrahmens verlässt, kann man erkennen, dass ein Schöpfer die Feder ohne Probleme erklären kann.
Und im Gegensatz zur Makroevolution – die weder im Labor nachgestellt werden konnte noch in der Natur beobachtet oder in der Fossilabfolge eindeutig nachgewiesen wurde – haben wir bei einer hypothetischen Schöpfung eine Vergleichsoption: Uns selber. Der Mensch ist selber schöpferisch tätig. Typisch für menschliche Erfindungen (Uhren, Motoren, Computer, u.v.m.) sind Feinabstimmung, Komplexität, Zukunftsplanung und noch mehr.
Die gleichen Merkmale sehen wir in oft noch viel stärkerer Form in der Natur, was einen Schluss auf einen Schöpfer mehr als nur rechtfertigt.
Ausführlicher wird die Entstehung der Feder und des Vogelfluges in diesem Artikel behandelt: https://www.wort-und-wissen.org/wp-content/uploads/b-17-1_feder-und-flug.pdf